
A. Reynoutria japonica Houtt. – křídlatka japonská (Fallopia japonica var. compacta a Fallopia japonica var. japonica)
B. Reynoutria sachalinensis (F. Schmidt) Ronse Decraene – křídlatka sachalinská
C. Reynoutria × bohemica (Chrtek & Chrtková) J. P. Bailey – křídlatka česká
Anglicky: A. Japanese knotweed, B. Giant knotweed, C. Bohemian knotweed
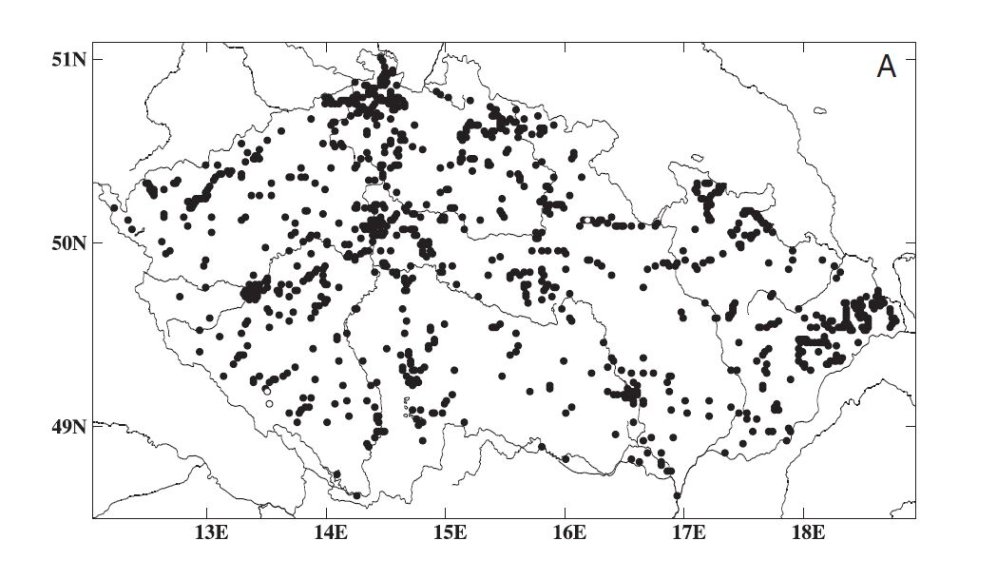
Obr. 1. Renutria japonica rozšíření v ČR (podle Mandáka a kol., 2004) 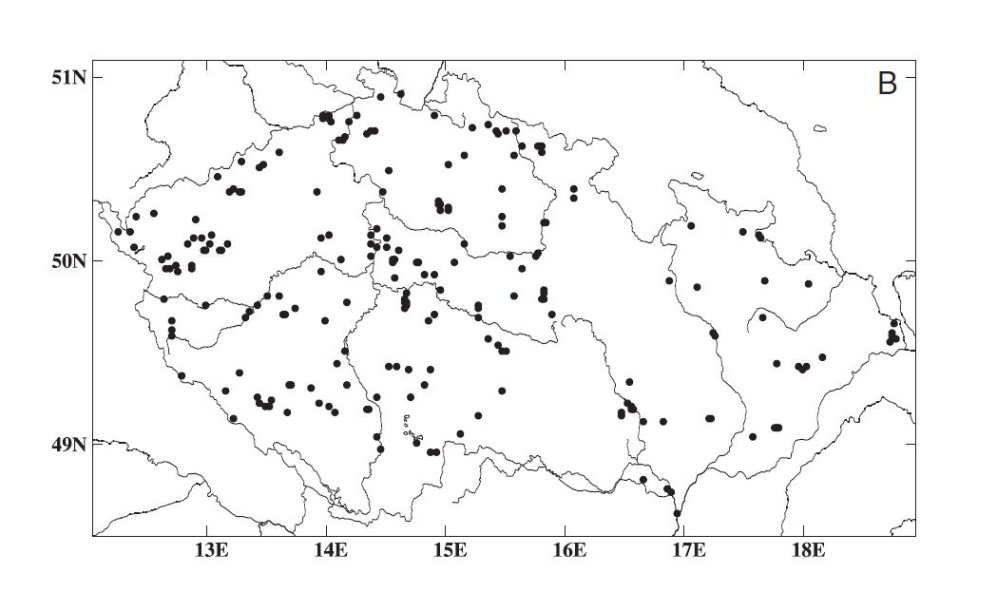
Obr. 2. Reynoutria sachalinensis rozšíření v ČR (podle Mandáka a kol., 2004) 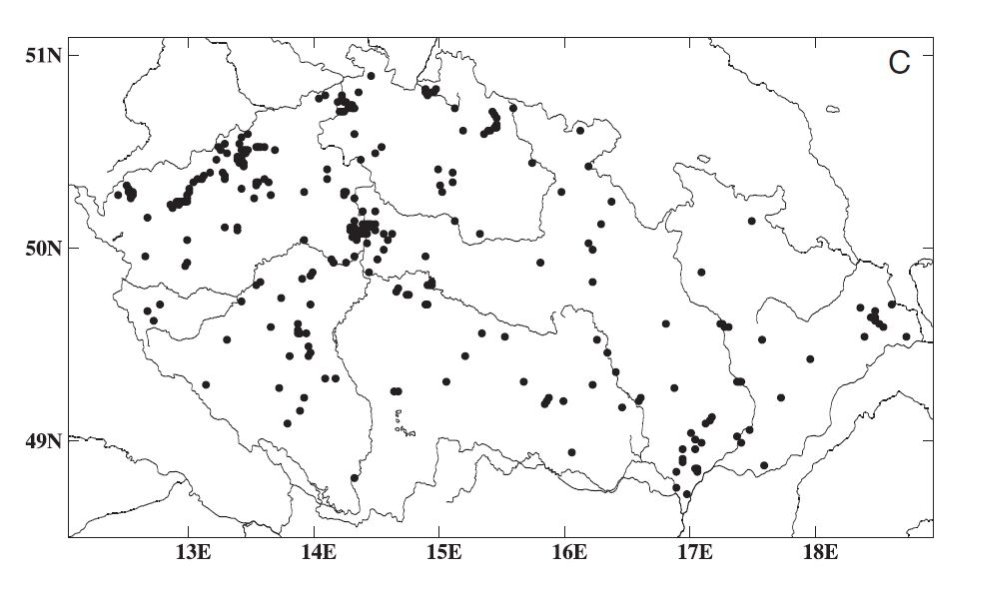
Obr. 3. Reynoutria x bohemica rozšíření v ČR (podle Mandáka a kol., 2004)
Synonyma:
A. Pleuropterus cuspidatus (Sieb. & Zucc.) Mildenke, Pleuropterus zuccarinii (Small) Small, Polygonum cuspidatum Sieb. & Zucc., Polygonum reynoutria Makino, Polygonum seiboldii Vriese, Polygonum zaccharini Small, Fallopia japonica Houtt., Tiniaria cuspidata (Houtt.) Hedb., Tiniaria japonica (Houtt.) Hedberg
B. Pleuropterus sachalinensis (F. Schmidt) H. Gross, Polygonum sachalinense F. Schmidt, Fallopia sachalinensis (F. Schmidt) Nakai, Reynoutria vivax J. Schmitz and Strank, Tiniaria sachalinensis (F.W. Schmidt ex Maxim.) Janch.
C. Polygonum x bohemicum (J. Chrtek & A. Chrtková) P.F. Zika & A.L. Jacobson, Polygonum x vivax J. Schmitz & Strank, Fallopia x bohemica J. Chrtek & A. Chrtková, Reynoutria x vivax J. Schmitz & Strank
Taxonomie:
oddělení Magnoliophyta (krytosemenné rostliny) » třída Rosopsida (vyšší dvouděložné) » řád Polygonales (rdesnotvaré) » čeleď Polygonaceae (rdesnovité)
Etymologie:
Rod Reynoutria byl pojmenován na počest vlámského mecenáše botaniky Karla van Sint Omaars (van Reynoutre). Rod Fallopia vychází z latinského slova fallo=klamu (Šmíd, 2002).
Ellenbergovy indikační hodnoty:
Reynoutria japonica
| Ellenbergovy indikační hodnoty (?) | |
|---|---|
| světlo | 88 |
| teplo | 66 |
| kontinentalita | 22 |
| vlhkost | 88 |
| půdní reakce | 55 |
| živiny | 77 |
Reynoutria sachalinensis
| Ellenbergovy indikační hodnoty (?) | |
|---|---|
| světlo | 77 |
| teplo | 77 |
| kontinentalita | 22 |
| vlhkost | 88 |
| půdní reakce | 77 |
| živiny | 88 |
Status: Reynoutria japonica var. compacta je přechodně zavlečeným druhem. Reynoutria japonica var. japonica, Reynoutria sachalinensis a Reynoutria × bohemica jsou klasifikovány jako invazní neofyty, tvoří metapopulace a patří mezi 100 nejinvazivnějších druhů Evropy. V Černém a šedém seznamu ČR jsou křídlatky zařazeny do druhové skupiny BL1 (neofytní byliny s největší mírou škodlivosti).
Životní forma: Geofyt
Primární areál: Primárním areálem Reynoutria japonica je podle některých autorů Japonsko (Yamazaki, 1994), podle jiných (např. Chrtek, 1990; Beerling a kol., 1994) i Korejský poloostrov, Čína a Taiwan. V rámci primárního areálu je druh morfologicky velmi variabilní, takže jeho taxonomie není zcela jasná (Mandák a kol., 2004). Například Yamazaki (1994) na základě odlišné morfologie separuje čínskou a korejskou populaci jako Reynoutria forbesii, Yonekura a Ohashi (1997) vymezují kromě nominátní variety R. japonica var. japonica i další dvě – var. hachidyoensis a var. uzenensis, endemické v Japonsku, které nebyly nikdy introdukovány mimo svůj primární areál. Na rozdíl od primárního areálu je klon vyskytující se v Evropě cytologicky i geneticky jednotný (Mandák a kol., 2004). Zakrslá horská varieta F. japonica var. compacta, rostoucí na Mt. Fuji (Japonsko), není v primárním areálu rozlišována, protože velká morfologická variabilita zde vytváří kontinuum mezi R. japonica var. japonica a R. japonica var. compacta (Shiosaka a Shibata, 1993).
Primárním areálem Reynoutria sachalinensis je Japonsko (severní Honshu a Hokkaido), ostrov Sachalin (Rusko) a možná i izolovaný ostrov Ullung-do (mezi Japonskem a Koreou; Bailey a Conolly, 2000). Yonekura a Ohashi (1997) rozlišují R. sachalinensis var. intermedia, rostoucí na severu ostrova Hokkaido. Vztah k Reynoutria × bohemica nebyl nikdy studován, ani na místech přirozeného výskytu obou rodičů tohoto křížence.
Výskyt Reynoutria × bohemica, křížence mezi R. japonica and R. sachalinensis, byl z primárního areálu překvapivě potvrzen relativně nedávno jako Reynoutria × mizushimae (Mandák a kol., 2004). Pravděpodobně se vyskytuje v místech překryvu areálů obou rodičovských druhů, tedy na japonských ostrovech Hokkaido a Honshu.
Reynoutria japonica vyhledává slunná místa v pahorkatinách a horách, kolonizuje široké spektrum stanovišť – roste na ruderálních místech, na silničních okrajích a v příkopech, štěrkových březích řek, na vychládajících lávových polích v alpínských polohách i obhospodařovaných pastvinách, kde se stává obtížným plevelem. Je rovněž součástí okrajů lužních lesů nebo lesů s dominancí Quercus serrata (např. Makita a kol., 1979; Hirose a Tateno, 1984; Bailey, 2003). V Mt. Fuji se F. japonica vyskytuje nad horní hranicí lesa v nadmořských výškách 2500-2600 m, což je její výškové maximum (Maruta 1983). Reynoutria sachalinensis vytváří téměř neproniknutelné porosty na erodovaných březích potoků a řek, v Japonsku až do nadmořské výšky 1050 m. Reynoutria × bohemica se pravděpodobně stejně jako rodičovské druhy vyskytuje na synantropních stanovištích a podél vodních toků.
Historie introdukce a šíření: Reynoutria japonica a R. sachalinensis byly introdukovány do Evropy jako okrasné rostliny v 19. století a brzy se začaly spontánně šířit do okolí. Historií šíření obou druhů se na Britských ostrovech zabývá Conolly (1977), v ostatních oblastech Jäger (1995). První žijící rostlina křídlatky japonské byla dovezena v roce 1825 z Číny a byla pěstována v londýnském parku Chiswick, ale uhynula. V Evropě rostoucí křídlatka pochází pravděpodobně z jediného klonu, dovezeného koncem 40. let 19. století Philippem von Sieboldem do města Leiden v Nizozemsku, odkud se jako Polygonum sieboldii prodávala ve formě oddenků do celé Evropy (Bailey a Conolly, 2000). Stala se oblíbeným prvkem viktoriánských zahrad, vychvalován byl její rychlý růst a krása, zdraví prospěšné účinky oddenků i možnost použití mladých listů místo zeleniny. Pěstována byla rovněž jako rychle rostoucí a pro dobytek chutná pícnina nebo jako medonosná rostlina. Zemědělci ji vysazovali pro ochranu svých choulostivých kultur před prudkým větrem a ostrým sluncem, nebo jako protierozní rostlinu (Jäger 1995; Bailey a Conolly, 2000). Už v roce 1847 byla v Utrechtu vyhlášena za nejzajímavější novou rostlinu roku. V roce 1841 uvedl katalog společnosti Von Siebold&Company na trh zakrslou horskou variantu křídlatky japonské – Polygonum pictum (neplatné synonymum od Reynoutria japonica var. compacta), která však i přes časnou introdukci nikdy nedosáhla ani popularity, ani rozšíření jako nominátní varieta (Mandák a kol. 2004).
Nejstarší herbářová položka Reynoutria japonica var. japonica byla pořízena Antonínem Weidmannem v roce 1883 v parku v Netolicích. Ve volné krajině byla tato křídlatka poprvé zaznamenána už v roce 1902 J. Soukupem v silničním příkopu u zahrady v Jilemnici. Na začátku 20. století zplaňovala jen výjimečně, zlom však nastal v meziválečném období, kdy byla běžně nabízena v zahradnictvích. Docházelo k únikům ze zahrad a parků (Obr. 4), kde byly křídlatky populární pro jejich rychlý růst jako krycí dekorativní rostliny, působivé zejména ve velkých porostech. Do krajiny se začala exponenciálně šířit dřív Reynoutria japonica – po 46 letech od introdukce než R. sachalinensis – po 83 letech (Pyšek a Prach, 1993). Do roku 2000 bylo z celkového počtu 1982 lokalit křídlatek rostoucích na území ČR zaznamenáno 1335 lokalit křídlatky japonské pravé (Reynoutria japonica var. japonica), a to na širokém spektru stanovišť (Mandák a kol., 2004). Historický nárůst počtu lokalit ukazují záznamy z Nálezové databáze AOPK ČR.
Na území ČR vzácná křídlatka japonská tuhá (Reynoutria japonica var. compacta) byla botaniky dlouho přehlížena. Přestože první doložený záznam o jejím pěstování pochází ze zahrady u Rychnova nad Kněžnou už z roku 1948, zplanělý byl tento taxon nalezen až v roce 1995 na Šumavě (Mandák a kol., 2004).
Křídlatka sachalinská (Reynoutria sachalinensis) byla do Evropy introdukována hned třikrát – H. Weyrichem v roce 1855, poté F. Schmidtem v roce 1861 a C. J. Maximoviczem v roce 1864. Ze Sachalinu byl rostlinný materiál převezen do botanické zahrady v Petrohradě a odtud distribuován do celého světa (Bailey a Conolly, 2000). Na území ČR byl tento druh poprvé sbírán V. Vlachem v roce 1921 v Dolních Krutech u Kolína, historické záznamy však podobnou rostlinu zmiňují už z roku 1869 v Třeboni (Pyšek a Prach, 1993). Historický nárůst počtu lokalit křídlatky sachalinské ukazují záznamy z Nálezové databáze AOPK ČR.
Výskyt křížence Reynoutria japonica var. japonica a R. sachalinensis, tj. R. × bohemica v severním Japonsku byl překvapivě potvrzen teprve nedávno (Bailey, 2003). Neexistují o něm spolehlivé historické záznamy. Nejstarší herbářová položka pochází z botanické zahrady v Manchesteru; v roce 1872 byl snad pěstován v anglických botanických zahradách. Bailey a Conolly (2000) také zmiňují druh “Polygonum Cooki ”, který byl pravděpodobně introdukován ze Severní Ameriky, nebo vznikl hybridizací až v Británii. Podle nepříliš kvalitní fotografie by se mohlo jednat o křídlatku českou nebo sachalinskou. První herbářový sběr křížence z území ČR pochází od Josefa Dostála, a to z Botanické zahrady University Karlovy v Praze, kde byl v roce 1950 pěstován. V roce 1983 byl tento taxon z našeho území (z Náchodska) popsán. Přestože je první doklad o výskytu tohoto hybrida poměrně nedávného data, jedná se o taxon v ČR velmi běžný. V minulosti byl mylně determinován jako R. sachalinensis, jen s menšími listy. Historický nárůst počtu lokalit křídlatky české ukazují záznamy z Nálezové databáze AOPK ČR.

Obr. 4. Z výsadeb v chatových koloniích se křídlatky mohou šířit do okolní krajiny (Sokolovsko). 
Obr. 5. Křídlatky mají statné, až 4 m vysoké, přímé lodyhy, v horní části větvené, silné a duté.
Popis druhu a možné záměny: Křídlatky jsou vytrvalé dvoudomé byliny s bohatě rozvětvenými, silnými dlouhými oddenky. Lodyhy jsou statné, až 4 m vysoké (Reynoutria sachalinensis; R. japonica maximálně 2 m), přímé, v horní části větvené, silné a duté (Obr. 5).
Křídlatka japonská (Reynoutria japonica) má menší listy (5-17 cm dlouhé a 5-12 cm široké) s okrouhlou až široce trojúhelníkovitou čepelí, na bázi kolmo uťatou. Listy jsou na rubu nezřetelně chlupaté, lupou lze pozorovat krátké papily se silně nafouklou bází. Křídla okvětí jsou široká, po květní stopce nesbíhavá. Nominátní varieta Reynoutria japonica var. japonica je vyšší (až 2,5 m) s většími listy (10-17 x 8-12 cm), široce trojúhelníkovitými, u horních listů zakončenými dlouhou úzkou špičkou. Kvete bíle. Reynoutria japonica var. compacta je max. 1,3 m vysoká, s menšími listy, v obrysu téměř okrouhlými, u horních listů zakončenými tupou trojúhelníkovitou špičkou. Křídla okvětí jsou narůžovělá, za plodu vínově červená (Kubát, 2002).
Křídlatka sachalinská (Reynoutria sachalinensis) bývá v průměru větší, má větší listy (20-30 cm dlouhé a 10-20 cm široké) s podlouhle vejčitou čepelí, na vrcholu zaokrouhlenou a na bázi hluboce srdčitou. Listy jsou na rubu roztroušeně dlouze chlupaté stejně dlouhými chlupy, na bázi neztlustlými. Křídla okvětí jsou úzká, po květní stopce výrazně sbíhavá (Chrtek, 1990).
Křídlatku českou (Reynoutria × bohemica) lze snadno zaměnit za rodičovské druhy, nejpodobnější je samčím rostlinám křídlatky sachalinské, jejíž květy záhy po odkvětu opadávají. Křídlatka česká je jen zřídka plodná. Rozdíl mezi jednotlivými druhy křídlatek je názorně prezentován na stránce Květeny ČR nebo NOBANIS.
Křídlatky kvetou od července do září. Květenstvím je lata mnohokvětých lichoklasů, květy nemají rozlišen kalich a korunu. U křídlatky japonské jsou okvětní lístky bílé, vzácněji slabě narůžovělé, u křídlatky sachalinské zelenobílé, vzácně žlutavé. Plodem je 3-hranná nažka.
Ekologie: Křídlatky preferují půdy s větším obsahem živin, mírně kyselé a vlhké. Patří k hemiheliofytům. Za nejdůležitější faktor ovlivňující jejich výskyt je považováno pravidelné narušování stanovišť (Bímová a kol., 2003), proto jsou vázány na lidská sídla a nejčastěji rostou na březích vodních toků (Obr. 7, 8, 9, 11, 13), podél silnic (Obr. 5, 10, 12) a železnic. Najdeme je také na rumištích (Obr. 6), lesních okrajích, ojediněle ve světlých lesích, na loukách a pastvinách, výjimečně na orné půdě (Beerling a kol., 1994). Právě management jako pastva, pravidelná seč nebo orba křídlatky výrazně potlačuje. Záměrně vysazeny bývají v zahradách, parcích nebo jako živé ploty.
Křídlatky vytvářejí na invadovaných stanovištích husté, zapojené porosty, které zcela potlačují ostatní vegetaci (Obr. 13). Velkou hrozbou jsou zejména pro společenstva aluvií řek, kde dochází k téměř úplné eliminaci ostatních druhů. Vedle křídlatek se vyskytují pouze jarní geofyty, které jsou adaptovány na růst ve specifických podmínkách opadavých lesů (např. Ficaria bulbifera), nebo ruderální nitrofyty jako např. Urtica dioica, Geranium robertianum, nebo Aegopodium podagraria. Nejméně invadovaná jsou sukcesně pokročilejší společenstva (lesy) a biotopy s pravidelným managementem, který obecně omezuje růst křídlatek (louky, pole, sekané plochy předzahrádek; Bímová a kol., 2004). Bímová a kol. (2004) zjistili, že druhová bohatost postiženého společenstva nemá vliv na úspěšnost invaze křídlatkami. Důležitější je kombinace stanovištních podmínek a počet propagulí (tj. částí organismu sloužící k šíření a reprodukci).
Reynoutria japonica je u nás často pěstována jako dekorativní rostlina v parcích a zahradách, odkud zplaňuje na synantropní i přirozená stanoviště. Invaduje více stanovišť než ostatní křídlatky. Je nenáročná na substrát, roste na všech typech půd, včetně vysychavých a rašelin, dokáže tolerovat nedostatek dusíku (Beerling a kol., 1994) i vysoké koncentrace těžkých kovů (Kubota a kol., 1988). Rovněž snáší nadlimitní hodnoty oxidu siřičitého v městském prostředí (Beerling a kol., 1994). Podle Beerlinga a kol. (1994) je omezena klimaticky – limitují ji roční úhrny srážek nižší než 500 mm a nedostatek letních srážek, což vysvětluje její absenci v kontinentální východní Evropě nebo subtropických oblastech. Dále je citlivá na jarní a podzimní mrazy. Křídlatka japonská je silně invazní druh, šíří se pomocí úlomků oddenků nebo lodyh zejména podél vodních toků (hlavně na erodovaných březích, Obr. 7-9) a komunikací, na skládkách, rumištích, opuštěných plochách, nebo na okrajích vlhkých křovin. Jasná je vazba na narušená stanoviště a městské prostředí, stejně jako na dopravní koridory (Obr. 12). Její regenerační schopnost je velmi vysoká, stačí oddenek o váze menší než 0,7 g, který je vodou nebo činností člověka zanášen na dlouhé vzdálenosti. Následně dojde k regeneraci rostliny z těchto fragmentů. Spolu s křídlatkou českou regeneruje křídlatka japonská až 2x lépe z oddenku než z lodyhy, zatímco křídlatka sachalinská regeneruje lépe z lodyhy (Bímová a kol., 2003). Generativní rozmnožování není možné, protože do Evropy byl zavlečen pouze jediný samičí klon a žádný samčí (Mandák a kol., 2005), což potvrzuje i zjištění jediného genotypu tohoto druhu na území ČR (Pyšek a kol., 2003). Přestože se křídlatka japonská nemůže generativně rozmnožovat, bývají rostliny na podzim bohatě plodné. Plody většinou vznikají v důsledku opylení druhem Reynoutria aubertii (Kubát, 2002). Semenáčky jsou ve volné přírodě velmi vzácné. Křídlatka japonská dává přednost kyselým a vlhkých půdám. Vyhledává otevřená stanoviště, zastínění snižuje její vzrůst i pokryvnost, proto není schopna invadovat do lesních porostů (např. Beerling, 1991).
V sekundárním areálu se vyskytují jak funkčně samičí, tak i funkčně samčí rostliny Reynoutria sachalinensis. K pohlavnímu rozmnožování v rámci tohoto druhu nepravidelně dochází, o čemž svědčí i genetická variabilita, která je ve srovnání s předchozím druhem mnohem vyšší (Mandák a kol., 2005). Křídlatka sachalinská je často pěstována v parcích a zahradách, ochotně zplaňuje na březích vodních toků, okrajích vlhkých křovin a v intravilánu obcí. Nová území invaduje hlavně vegetativně, dopravou, přenosem kontaminované půdy, štěrku nebo písku a proudící vodou (ideálně při záplavách). Je méně invazní než ostatní dvě křídlatky, vyznačuje se menší schopností regenerace z lodyhy i oddenků a obvykle netvoří tak rozsáhlé porosty na březích vodních toků (Bímová a kol., 2003).
Reynoutria × bohemica s nejvyšší pravděpodobností vzniká přímo na lokalitách v ČR, což bylo prokázano na základě isozymové analýzy (Mandák a kol., 2005). Zavlečení křížence z míst původního areálu, kde dochází ke styku rodičovských druhů, není pravděpodobné.
Křídlatka česká se sice šíří zejména vegetativní cestou, tj. regenerací z fragmentů oddenků a lodyh, Bímová a kol. (2004) ale zjistili, že občas může vzniknout i ze semen, pokud spolu na lokalitě roste samičí rostlina Reynoutria japonica var. japonica (samčí klon se v Evropě nevyskytuje) a fertilní samčí Reynoutria sachalinensis. Tak jako ostatním křídlatkám i křídlatce české vyhovují vlhčí, živinami bohaté půdy s pravidelným mechanickým narušováním svrchní vrstvy. Nejčastěji se vyskytuje na místech alespoň v minulosti narušených lidskou činností, ale stále častěji expanduje i do ryze přírodních společenstev, zejména podél potoků a řek (Chrtek, 1990). Má lepší regenerační schopnosti než rodiče, je konkurenčně schopnější, takže je často z lokalit společného výskytu vytlačuje (Bímová a kol., 2003).

Obr.6. Křídlatky se snadno šíří pomocí úlomků oddenků nebo lodyh. S odpadem ze zahrádek se tak dostávají na skládky, kde se mohou uchytit a dále šířit (skládka Ředhošť). 
Obr. 7. Rezavé podzimní zbarvení křídlatek je v některých břehových porostech velmi výrazné (údolí Bečvy). 
Obr. 8. Pro rozšíření křídlatek je důležité pravidelné narušování stanoviště, proto jim vyhovují břehy vodních toků (souvislé porosty v údolí Ohře).
Vegetace: Ve Vegetaci ČR (Chytrý, 2009) jsou porosty křídlatek zařazeny do samostatné asociace Reynoutrietum japonicae Görs et Müller in Görs 1975 v rámci svazu Aegopodion podagrariae. Jsou vyčleněny tři varianty podle dominantního druhu: (1) varianta Reynoutria japonica, (2) varianta R. sachalinensis a (3) varianta R. x bohemica.
Rozšíření: Reynoutria sachalinensis byla introdukována a je vnímána jako vysoce invazní a značně nebezpečný druh napříč Evropou i Severní Amerikou, v Austrálii i na Novém Zélandě a dále byla zaznamenána v Indii a Jihoafrické republice. R. japonica v současnosti roste především v severní a střední Evropě, až do 68° s. š. Zaznamenána byla i v jižní Evropě, jako např. v Chorvatsku, Makedonii, Bosně a Hercegovině (Trinajstic, 1990). Invazní je i v různých částech Severní Ameriky, Austrálie a Nového Zélandu, zavlečena byla i do Jižní Ameriky (Chile). R. x bohemica je invazní na většině území Evropy a v Severní Americe, v jižnějších a východnějších částech Evropy je zřejmě hojnější než rodičovské druhy. Rozšíření křídlatek v Evropě podrobně uvádí DAISIE – R. japonica, R. sachalinensis a R. x bohemica.
V České republice jsou křídlatky rozšířeny po celém území ČR od nížin až do podhorského stupně (Chrtek, 1990). Těžiště je v mezofytiku, hojné jsou i v termofytiku; v oreofytiku zplaňují jen v nižších polohách, kde je místy častější Reynoutria sachalinensis než R. japonica. Křídlatka sachalinská se v posledních letech šíří rychleji než dřív, na některých lokalitách vytváří rozsáhlé porosty (např. u Jizery nedaleko Mladé Boleslavi, v okolí Děčína), přesto je její rychlost šíření stále nižší než u křídlatky japonské. Křídlatka česká roste spolu s rodičovskými druhy i zcela odděleně. Ve srovnání s rodičovskými druhy se šíří dvojnásobnou rychlostí (Mandák a kol., 2004). Ohromujících rozměrů dosahují její porosty v místech, kde se chová jako hybridogenní taxon (např. Severní Morava), takže ke svému vzniku nepotřebuje opakovanou hybridizaci rodičovských druhů, ale dochází k opylování a generativnímu rozmnožování přímo v rámci křížence (Mandák a kol., 2005).
Křídlatky se nejčastěji vyskytují v blízkosti lidských sídel, což souvisí s historií jejich introdukce, kdy byly pěstovány jako okrasné rostliny ve městech i vesnicích a odtud se činností člověka, vody nebo transportem půdy při stavebních pracích rozšířily do volné krajiny. Nejmenší afinitu k osídlení vykazuje R. x bohemica, která preferuje břehové porosty podél vodních toků. R. japonica var. japonica upřednostňuje okolí silnic, zatímco R. sachalinensis je ze všech tří druhů nejčastější součástí zahrad a parků (Mandák a kol., 2004).
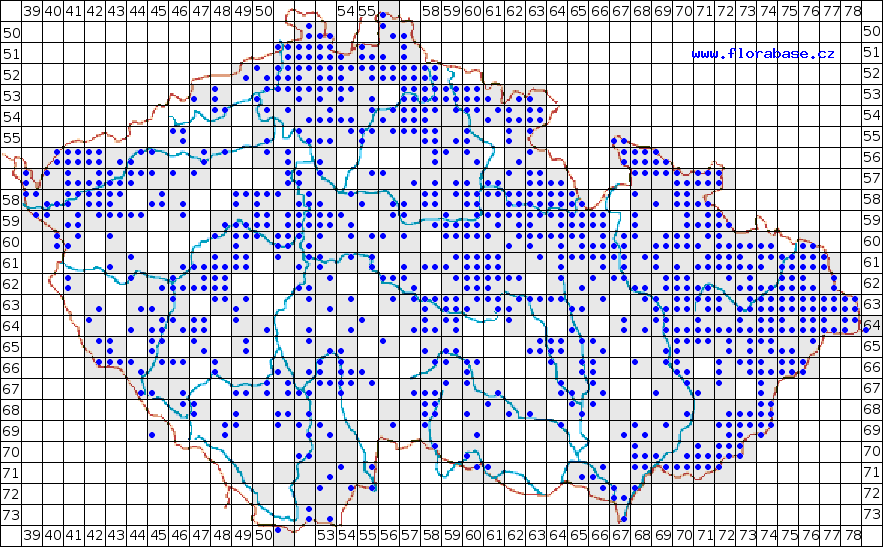
Na území ČR je nejhojnější nejdříve introdukovaná R. japonica var. japonica (Obr. 1), která se podle Mandáka a kol. (2004) v roce 2000 vyskytovala na 1335 lokalitách. R. x bohemica (Obr. 3)byla zjištěna na 381 lokalitách, R. sachalinensis (Obr. 2) na 261 lokalitách a R. japonica var. compacta na 5 lokalitách, z toho 3 jsou zplanělé.
Křídlatky se nejčastěji vyskytují v blízkosti lidských sídel, což souvisí s historií jejich introdukce, kdy byly pěstovány jako okrasné rostliny ve městech i vesnicích a odtud se činností člověka, vody nebo transportem půdy při stavebních pracích rozšířily do volné krajiny. Nejmenší afinitu k osídlení vykazuje R. x bohemica, která preferuje břehové porosty podél vodních toků. R. japonica var. japonica upřednostňuje okolí silnic, zatímco R. sachalinensis je ze všech tří druhů nejčastější součástí zahrad a parků (Mandák a kol., 2004).
Využití: Křídlatky se hojně používaly (a stále je lze najít v nabídce internetových obchodů) jako významné dekorativní, krycí parkové rostliny, působivé zejména v rozsáhlých porostech. Mladé rostliny poskytující velké množství rychle rostoucí rostlinné biomasy, jsou vhodné jako čerstvé krmivo pro dobytek, nikoliv ale v podobě sena. Kvůli rychlému růstu byly křídlatky vybrány pro pěstování v energetických plantážích a na výrobu biopaliva. Křídlatky se také používají jako erozní a stabilizační rostlina, k rekultivacím skládek a výsypek, nebo k asanacím půdy kontaminované těžkými kovy. Oddenky křídlatek jsou používány v tradiční asijské medicíně. Květy i olistěné lodyhy jsou používány k přípravě kulinářských specialit.

Obr. 9. V údolí Ohře se křídlatky vyskytují spolu s bolševníkem. 
Obr. 10. Jasná je vazba křídlatek na dopravní koridory. 
Obr. 11. Křídlatky jsou často součástí břehových porostů. Snášejí i částečné zastínění ve vrbinách nebo olšinách (dolní Ohře).
Management: Křídlatky jsou napříč Evropou i v řadě zemí Severní Ameriky na seznamech nebezpečných invazních druhů, jsou na seznamu karanténních rostlinných škůdců Q-banky i na seznamu invazních druhů, které nejvíce ovlivňují biodiverzitu v Evropě, mají negativní vliv na lidské aktivity, zdraví i ekonomické zájmy. Legislativa řešící jednotně na úrovni celé Evropské unie management takto škodlivých invazních druhů se teprve zpracovává, jednotlivé evropské státy proto zatím řeší tuto problematiku individuálně. Ve Velké Británii, Španělsku (jen R. japonica), Slovensku, Polsku a Estonsku se zakázán dovoz, pěstování, prodej nebo záměrné šíření křídlatek do volné krajiny. V České republice v současné době neexistuje žádné přímé legislativní nařízení, které by upravovalo nakládání s křídlatkami. V zákoně o ochraně přírody a krajiny č. 114/1992 Sb. je pouze ustanoveno, že záměrné šíření geograficky nepůvodního druhu rostliny nebo živočicha je možné jen s povolením orgánu ochrany přírody. V minulosti křídlatky podobně jako bolševník podléhaly monitoringu Státní rostlinolékařské správy, v aktuálním prováděcím předpisu k rostlinolékařskému zákonu, tj. ve vyhlášce č. 215/2008 Sb. v platném znění, již nejsou zařazeny.
Protože jsou všechny křídlatky konkurenčně velmi silné, s vysokou schopností šířit se, jsou podle Černého a šedého seznamu tolerovány pouze v suburbánních oblastech. Ve všech ostatních případech (např. kultury, spontánní populace a metapopulace v intravilánech i ve volné krajině) musí být likvidovány a nové porosty by neměly být zakládány. Metody likvidace křídlatek testoval Barták a kol. (2010) v Moravskoslezském kraji v rámci projektu LIFE, hojně je používaný tzv. Beskydský postup (Šrubař a Albín, 2005). Z výsledků těchto projektů vychází i standard „Likvidace vybraných nepůvodních druhů rostlin“ (Pergl a kol., 2014).
Protože se křídlatky rozmnožují téměř výhradně vegetativně a vytvářejí mohutný podzemní kořenový systém, je jejich likvidace obtížná a velmi zdlouhavá. Vzhledem k vysoké schopnosti regenerovat z velmi malých úlomků oddenků i nadzemních výhonů vyžaduje management pečlivý přístup a opatrné nakládání s rostlinným odpadem (posekanou biomasou) a zeminou kontaminovanou oddenky křídlatek. Při zemních pracích je na lokalitách s výskytem křídlatek třeba ukládat kontaminovanou zeminu zvlášť a zabránit tak jejich rozšíření na celou plochu. Nejvhodnější metodou likvidace křídlatek je aplikace herbicidu na list. Při managementu musí být herbicidem zasažena maximální listová plocha. U rozsáhlejších porostů není možné omezit aplikaci postřikem „shora“ a do porostů je nutné prosekat průchody, aby bylo možné herbicid aplikovat i na spodní patra listů. Průseky je pak třeba ošetřit po jejich zmlazení nebo v následujícím roce. Nejvyšší účinnost v poměru k vynaloženým nákladům vykazuje aplikace herbicidu v pozdní vegetační sezóně (koncem léta). Injektáž je vhodná na ochranářsky hodnotných lokalitách, kde hrozí riziko zasažení okolní vegetace herbicidem, nebo v okolí vodních zdrojů a v jejich ochranných pásmech. Bodová aplikace herbicidu vpichy do stvolů nebo nátěr stvolů je metoda velmi šetrná k okolní vegetaci, minimalizuje i zasažení vody a půdy. Injektáž lze aplikovat i za horšího počasí (vítr, slabý déšť). Metoda je sice velice účinná již po první sezóně, pokud se ale v následujícím roce objeví regenerující rostliny, které nelze injektovat vzhledem k jejich slabému vzrůstu, použije se ošetření herbicidem na list. Roundup Biaktiv se používá v 20–30% koncentraci, aplikační dávka herbicidu je volena v rozmezí 3–7 ml do jednoho stvolu křídlatky. Injektáž je nutné aplikovat do cca 50 % stvolů v polykormonu. Minimální doporučená výška křídlatek pro použití této metody je 1,5 m a průměr stonků minimálně 1,5 cm (nejčastěji 1,5–5 cm). Zároveň je za stejných podmínek možno použít nátěr na list a stvol.
Přestože mechanické metody (pastva, sekání) nevedou k totální likvidaci křídlatek, jsou používány pro omezení jejich negativního vlivu na společenstva nebo na plochách, kde z různých důvodů nelze použít herbicidy (Obr. 13). Obtížně přístupné lokality se sekají křovinořezy, mačetami a kosami. Z důvodu rychlého růstu a dřevnatění stvolů se ruční kosení doporučuje opakovat několikrát za rok a vždy co nejníže u země. Při optimální výšce rostlin 40 cm je možné kosení až 8 krát ročně. Tato metoda účinně brání rozrůstání křídlatek, jen pokud je aplikována opakovaně a dlouhodobě. Na lokalitách s možností pastvy lze křídlatky omezovat pastvou a dosekáváním. Pastva musí být zahájena co nejdříve kvůli stravitelnosti křídlatky dobytkem.
Vyrývání a vykopávání křídlatek se nedoporučuje z důvodu vysoké regenerační schopnosti z úlomků kořenů a finanční náročnosti. Metoda je velmi pracná a z hlediska dalšího šíření těchto rostlin riziková. Úspěšná je pouze v počátečním stádiu výskytu křídlatek nebo při výskytu jednotlivých rostlin ve velmi malých populacích. Vykopány musí být celé rostliny včetně oddenků (rhizomů), které mohou být zakořeněny až do hloubky 2 m. Minimální hloubka vyrývání je 30 cm. Vytrhaná a vykopaná biomasa se musí usušit a spálit. Zásah musí být proveden opakovaně v průběhu vegetační sezóny, vždy po obnově porostu. Je nutné dbát zvýšené opatrnosti a minimalizovat přesun (převoz) rostlinného materiálu a kontaminované zeminy. Po ošetření lokalit musí následovat výsev vhodné travní směsi.
Analýza rizika: Křídlatky jsou díky rozsáhlému systému rozvětvených oddenků schopny enormního vegetativního rozrůstání, kdy vytvářejí veliké polykormony. Generativní šíření pomocí semen je vzácné, hlavní roli hraje vegetativní reprodukce pomocí malých úlomků oddenků i lodyh, které mohou být při povodních nebo činností člověka šířeny i na velké vzdálenosti. Díky rychlému růstu a tvorbě značného množství biomasy během krátké doby mají křídlatky velkou konkurenční schopnost, vytlačují původní vegetaci a vytvářejí neprostupné monocenotické porosty. Klonální růst jim umožňuje dobře regenerovat, takže se mohou na stanovišti udržet velmi dlouho. Klony R. japonica var. japonica v 85,7 % případů a klony R. sachalinensis v 75,4 % na invadovaném stanovišti přetrvávají a dosahují i velmi vysokého věku. U křídlatky japonské byl zjištěn minimální věk nejstarších přežívajících klonů 91 let, u křídlatky sachalinské dokonce 130 let (Pyšek a kol., 2001). Protože jsou všechny druhy křídlatek jednotně vnímány jako agresivní invazní druhy a perspektivně se předpokládá jejich další šíření, je s výjimkou suburbánních oblastí doporučena jejich likvidace, a to pokud možno ještě v počátku, než dojde ke vzniku souvislých porostů. Nové porosty, ať už z okrasných nebo hospodářských důvodů, by neměly být zakládány.

Užitečné odkazy:
- biolib.cz/cz/taxon/id38823/
- biolib.cz/cz/taxon/id38826/
- biolib.cz/cz/taxon/id38822/
- botanickafotogalerie.cz/
- botany.cz/cs/reynoutria-bohemica/
- botany.cz/cs/reynoutria-sachalinensis/
- botany.cz/cs/reynoutria-japonica/
- cabi.org/isc/datasheet/107744
- cabi.org/isc/datasheet/23875
- cabi.org/isc/datasheet/108332
- europe-aliens.org/speciesFactsheet.do?speciesId=8137
- europe-aliens.org/speciesFactsheet.do?speciesId=8170
- europe-aliens.org/speciesFactsheet.do?speciesId=8037
- fs.fed.us/database/feis/plants/forb/polspp/all.html
- invasive.org/browse/detail.cfm?imgnum=5472099
- invasive.org/browse/detail.cfm?imgnum=1196049
- invaznidruhy.nature.cz/res/data/154/020207.pdf
- issg.org/database/species/ecology.asp?si=91&fr=1&sts=sss&lang=EN
- portal.nature.cz/publik_syst/nd_nalez-public.php?idTaxon=39156
- portal.nature.cz/publik_syst/nd_nalez-public.php?idTaxon=39151
- portal.nature.cz/publik_syst/nd_nalez-public.php?idTaxon=39154
- plants.usda.gov/core/profile?symbol=POBO10
- plants.usda.gov/core/profile?symbol=POSA4
- sci.muni.cz/botany/vegsci/vegetace.php?lang=cz&typ=XDE10
- quick.florabase.cz/map/show/taxon/Reynoutria%20sachalinensis/callback/???
- quick.florabase.cz/map/show/taxon/Reynoutria%20japonica/callback/???¨
- quick.florabase.cz/map/show/taxon/Reynoutria%20x%20bohemica/callback/???

Literatura
- Bailey J. P. (2003): Japanese knotweed s.l. at home and abroad. In: Child L., Brock J. H., Brundu G., Prach K., Pyšek P., Wade P. M. & Williamson M. (eds.): Plant Invasions, Ecological Threats and Management Solutions. – Backhuys Publishers, Leiden: 183–196.
- Bailey J. P. & Conolly A. P. (2000): Prize-winners to paria hs: A history of Japanese Knotweed s. l. (Polygonaceae) in the British Isles. – Watsonia 23: 93–110.
- Barney J. N., Tharayil N., DiTommaso A. & Bhowmik P. C. (2006): The biology of invasive alien plants in Canada. 5. Polygonum cuspidatum Sieb. & Zucc. [=Fallopia japonica (Houtt.) Ronse Decr.]. – Canadian Journal of Plant Sciences 86: 887–905.
- Barták R., Konupková Kalousová Š. & Krupová B. (2010): Metodika likvidace invazních druhů křídlatek (Reynoutria spp.) – Morav.-slezs. kraj ve spolupráci s ČSOP Salamandr za fin. podpory EU: 1–34.
- Beerling D. J. (1991): The testing of cellular concrete revetment blocks resistant to growths of
- Reynoutria japonica Houtt. (Japanese knotweed). – Water Res Ox 25: 495–498.
- Beerling D. J., Bailey J. P. & Conolly A. P. (1994): Biological Flora of the British Isles No.183 Fallopia japonica. – J. Ecol. 82 (4): 959–979.
- Bímová K., Mandák B. & Pyšek P. (2003): Experimental study of vegetative regeneration in four invasive Reynoutria taxa (Polygonaceae). – Plant Ecology 166: 1–16.
- Bímová K., Mandák B. & Kašparová I. (2004): How does Reynoutria invasion fit the various theories of invasibility? – Journal of Vegetation Science 15: 495–504. Conolly A. P. (1977): The distribution and history in the British Isles of some alien species of
- Polygonum and Reynoutria. – Watsonia 11: 291–311.
- Hirose T. & Tateno M. (1984): Soil nitrogen patterns induced by colonization of Polygonum cuspidatum on Mt. Fuji. – Oecologia 61: 218–223.
- Child L, & Wade M. (2000): The Japanese knotweed manual. – Packard Publishing, Chichester, p 123.
- Chrtek J. (1990): Reynoutria Houtt., křídlatka. In: Hejný S., Slavík B., Hrouda L. & Skalický V. (eds.): Květena ČR 2. – Academia, Praha: 362–366.
- Chytrý M. (ed.) (2009): Vegetace České republiky 2. Ruderální, plevelová, skalní a suťová vegetace. Vegetation of the Czech Republic 2. Ruderal, Weed, Rock and Scree Vegetation. – Academia, Praha: 373–375.
- Jäger E. J. (1995): Die Gesamtareale von Reynoutria japonica Houtt. und R. sachalinensis (F. Schmidt) Nakai, ihre klimatische Interpretation und Daten zur Ausbreitungsgeschichte. –Schr.-R. f. Vegetationskde., Sukopp-Festschrift 27: 395–403.
- Kubát K., Hrouda L., Chrtek J. jun., Kaplan Z., Kirschner J. & Štěpánek J. [eds.] (2002): Klíč ke květeně České republiky. – Academia, Praha: 201–202.
- Kubota K., Nishizono H., Susuki S. & Ishii F. (1988): A Copper-Binding Protein in Root Cytoplasm of Polygonum cuspidatum Growing in a Matalliferous Habitat. – Plant and Cell Physiology 29: 1029–1033.
- Makita H., Miyagi T., Muira O. & Kikuchi T. (1979): A study of an alder forest and an elm forest with special reference to their geomorphological conditions in a small tributary basin. In: Miyawaki A. & Okuda S. (eds.): Vegetation und Landschaft Japans. – Bull. Yokohama Phytosociological Society Yokohama/Japan 16: 237–249.
- Mandák B., Pyšek P. & Bímová K. (2004): History of the invasion and distribution of Reynoutria taxa in the Czech Republic: a hybrid spreading faster than its parents. – Preslia 76: 15–64.
- Mandák B., Bímová K., Pyšek P., Štěpánek J. & Plačková I. (2005): Isoenzyme diversity in Reynoutria taxa: escape from sterility by hybridization. – Plant Systematics and Evolution 253: 219–230.
- Maruta E. (1983): Growth and survival of current-year seedlings of Polygonum cuspidatum at the upper distribution limit on Mt. Fuji. – Oecologia 60: 316–320.
- Pergl J., Perglová I., Vítková M., Pocová L., Janata T. & Šíma J. (2014): Likvidace vybraných nepůvodních druhů rostlin. – AOPK ČR a BÚ AVČR, Praha: 25 pp.
- Pergl J., Sádlo J., Petrusek A., Laštůvka Z., Musil J., Perglová I., Šanda R., Šefrová H., Šíma J., Vohralík V. & Pyšek P. (2016): Black, Grey and Watch Lists of alien species in the Czech Republic based on environmental impacts and management strategy. – NeoBiota 28: 1-37
- Pyšek P., Brock J. H., Bímová K., Mandák B., Jarošík V., Koukolíková I., Pergl J., Štěpánek J. (2003): Vegetative regeneration in invasive Reynoutria (Polygonaceae) taxa: the determinant of invasibility at the genotype level. – Amer J Bot 90: 1487–1495.
- Pyšek P., Mandák B., Francírková T. & Prach K. (2001): Persistence of stout clonal herbs as invaders in the landscape: a field test of historical records. In: Brundu G., Brock J., Camarda I., Child L. & Wade M. (eds.), Plant invasions: Species ecology and ecosystem management. – Backhuys Publishers, Leiden: 235–244.
- Pyšek P. & Prach K. (1993): Plant invasions and the role of riparian habitats: a comparison of four species alien to central Europe. – Journal of Biogeography 20: 413–420.
- Shiosaka H. & Shibata O. (1993): Morphological changes in Polygonum cuspidatum Sieb. et Zucc. reciprocally transplanted among different altitudes. – Jap. J. Ecol. 43: 31–37.
- Šmíd M. (2002): Průvodce odbornými názvy rostlin. Brázda, Praha: 320 pp.
- Šrubař M. & Albín R. (2005): Jak „beskydský postup“ likvidace křídlatek šetří nejen přírodu. Ochrana přírody 60 (3): 82–84.
- Trinajstic I. (1994): Prolog poznavanju rasprostranjenosti vrste Reynoutria japonica Houtt. (Polygonaceae) u Jugoslaviji. – Fragmenta herbologica Jugoslavic 19(2): 139–143.
- Yamazaki T. (1994): On Reynoutria yunnanensis (Lévl.) Nakai ex Migo. – J. Jap. Bot. 69: 179–180.
- Yonekura K. & Ohashi H. (1997): New combinations of East Asian species of Polygonum s. l. – J. Jap. Bot. 72: 154–161